|
|
 |
| PROGENESIS POSITIVE |
|
| PROGENESIS MODESTY MELINA GP-82-3YR-CAN |
|
|
|
|
| BACON-HILL PETY MODESTY |
|
| PROGENESIS DUKE MELEE VG-87-5YR-USA 9* |
|
|
|
|
| S-S-I MONTROSS DUKE |
|
| PEAK BOMBERO MAXIMA VG-87-6YR-USA DOM |
|
|
|
| TD TR TL TY MWT TV 99%-I |
HH1F HH2F HH3F HH4F HH5F HH6F HCDF HMWF |
| Num.Reg #: HOCANM13353510 |
aAa: 234165 |
DMS: 561,456 |
| Nacimiento: 02/04/2019 |
Kappa Caseina: AB |
Beta Caseina: A1A2 |
|
| PRODUCCION |
| 176 Rebaños |
813 Hijas |
97% Rep. |
|
MACE-G / 04-24 |
|
|
Leche lbs
2115 |
|
Grasa lbs
138 |
|
Grasa %
+0.19 |
|
Proteína lbs
75 |
|
Proteína %
+0.03 |
|
| NM$ 943 |
|
CM$ 951 |
|
FM$ 885 |
|
GM$ 854 |
|
DWP$ 843 |
|
Eficiencia de Conversiòn
344 |
|
IR
119 |
|
Comida Ahorrada
-8 |
|
Eficiencia Metano
108 |
|
|
|
|
Media de Producción
Leche 30,751 lbs
Grasa 1,312 lbs
Proteína 995 lbs
|
|
| SALUD Y REPRODUCCIÓN |
|
Immunity 97 |
|
|
|
|
|
|
|
|
|
| Fertilidad de las Hijas | -4.6 |
|
|
|
|
|
|
| Facilidad de Parto | 1.7% | 84% Rep. |
|
|
|
|
| Facilidad de Parto de las Hijas | 1.1% | 86% Rep. |
|
|
|
|
| Crías del Toro que Nacen Muertas | 5.3% | |
|
|
|
|
| Crías de las Hijas que Nacen Muertas | 4.5% | |
|
|
|
|
| TIPO |
| 122 Rebaños |
354 Hijas |
94% Rep. |
|
MACE / 04-24 |
|
|
|
| Estruct. y Capacidad | -0.72 | |
|
|
|
|
|
|
|
|
|
|
|
|
|
| Estatura |
|
|
Alta |
+0.71 |
| Fortaleza |
|
|
Débil |
-0.23 |
| Profundidad Corporal |
|
|
Profunda |
+0.35 |
| Estructura Lechera |
|
|
Cost. Abierta |
+1.74 |
| Ángulo de Grupa |
|
|
Isq. Bajos |
+1.90 |
| Anchura Grupa |
|
|
Ancha |
+0.40 |
| Patas Vista Lateral |
|
|
Curvas |
+1.98 |
| Patas Vista Posterior |
|
|
Cerradas |
-1.55 |
| Ángulo Podal |
|
|
Bajo |
-0.55 |
| Colocación de las Patas |
|
|
Atrás |
-0.13 |
| Inserción Anterior |
|
|
Fuerte |
+1.98 |
| Altura Inserción Posterior |
|
|
Alta |
+1.17 |
| Anchura Inserción Posterior |
|
|
Ancha |
+1.38 |
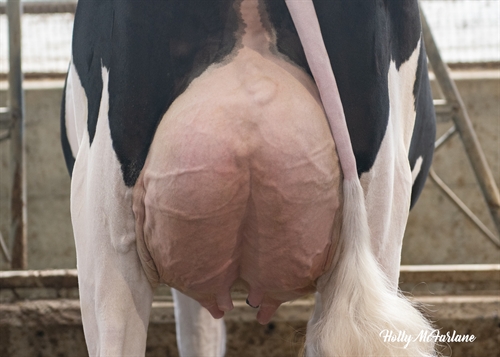
| Ligamento Suspensor |
|
|
Fuerte |
+0.38 |
| Profundidad Ubre |
|
|
Recogida |
+1.38 |
| Colocación Pezones Anteriores |
|
|
Cerrados |
+1.73 |
| Longitud de Pezones |
|
|
Cortos |
0.00 |
| Colocación Pezones Posteriores |
|
|
Cerrados |
+1.66 |
|
|
|
| |
|
|
|
|