|
|
 |
| MB-LUCKYLADY BULLSEYE |
|
| WESTCOAST ASHBY DOCA VG-85-2YR-CAN |
|
|
|
|
| ST GEN D-LAMBDA ASHBY |
|
| DICKLANDS KING DOC 3503 VG-87-4YR-CAN 2* |
|
|
|
|
| WOODCREST KING DOC |
|
| DICKLANDS CHIEF 2630 3041 VG-85-3YR-CAN |
|
|
|
| TD TR TL TY MWT TV 99%-I |
HH1F HH2F HH3F HH4F HH5C HH6F HCDF HMWF |
| Nom #: HOCANM14400215 |
aAa: 234165 |
DMS: 126,123 |
| Né le: 12/05/2022 |
Caséine kappa: BB |
Caséine bêta: A1A2 |
|
| PRODUCTION |
| G Troup |
G Filles |
82% Fiab |
|
CDCB-G / 04-26 |
|
|
Lait lbs
10 |
|
Gras lbs
21 |
|
Gras %
+0.08 |
|
Protéine lbs
4 |
|
Protéine %
+0.01 |
|
| NM$ -20 |
|
CM$ -15 |
|
FM$ -33 |
|
GM$ -67 |
|
DWP$ -244 |
|
Efficience de l'alimentation
27 |
|
IAR
14 |
|
Alimentation économisée
-147 |
|
Efficience du méthane
108 |
|
Vitesse de traite
6.76 |
|
| SANTÉ ET REPRODUCTION |
|
Immunité 95 |
|
|
|
|
|
|
|
| Taux de conception des vaches | -2.2 | |
|
|
|
|
| Taux de conception des génisses | 0.2 | |
|
|
|
|
| Facilité de vêlage | 1.8% | 86% Fiab |
|
|
| Livabilité des génisses | 0.7 |
|
|
| Facilité vêl. Filles | 1.9% | 71% Fiab |
|
|
|
|
|
|
|
|
| Mortinatalité des filles | 5.8% | |
|
|
|
|
| CONFORMATION |
| G Troup |
G Filles |
80% Fiab |
|
HAUSA-G / 04-26 |
|
|
|
|
| Stature |
|
|
Grande |
+2.39 |
| Force |
|
|
Forte |
+1.10 |
| Profondeur de corps |
|
|
Profond |
+2.00 |
| Car. Laitier |
|
|
Côtes ouvertes |
+2.17 |
| Angle de la croupe |
|
|
Isch. hauts |
-0.51 |
| Largeur de la croupe |
|
|
Large |
+1.95 |
| Mbres arr. Vue côté |
|
|
Courbés |
+1.75 |
| Mbres arr. Vue arr. |
|
|
Jarrets droits |
+1.93 |
| Angle du pied |
|
|
Profond |
+1.72 |
| Indice pieds & mbres |
|
|
Désirable |
+2.11 |
| Attache avant |
|
|
Forte |
+2.79 |
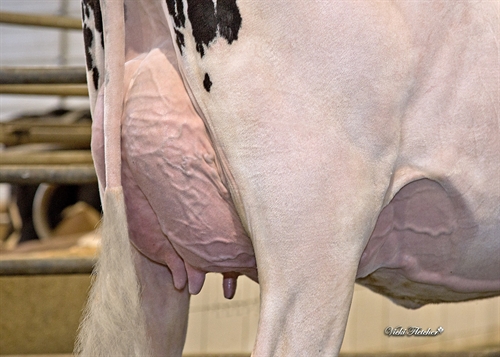
| Haut. Arrière-pis |
|
|
Haute |
+3.16 |
| Larg. Arrière-pis |
|
|
Large |
+3.07 |
| Suspension médiane |
|
|
Forte |
+2.22 |
| Profondeur du pis |
|
|
Haut |
+1.98 |
| Pos. trayons avant |
|
|
Rapprochés |
+1.70 |
| Longueur des trayons |
|
|
Courts |
-0.29 |
| Position trayons arrière |
|
|
Rapprochés |
+2.04 |
|
|
|
| |
|
|
|
|