|
|
 |
| VIEW-HOME POWERBALL-P |
|
| STANTONS MCCUTCHEN 1174 AGREE VG-88-4YR-CAN 17* |
|
|
|
|
| DE-SU BKM MCCUTCHEN 1174 |
|
| STANTONS OBSERVER EXTREME VG-85-2YR-CAN 11* |
|
|
|
|
| DE-SU OBSERVER |
|
| WABASH-WAY BOL ELIZABETH VG-87-2YR-USA DOM 19* |
|
|
|
| GLPI +3124 PRO$ 1341 |
| VG-CAN DPF MFF RDF BLF CNF BYF MWF CVF |
HH1F HH2F HH3F HH4F HH5F HH6F HCDF HMWF |
| Nom #: HO840M3129016082 |
aAa: 423516 |
DMS: 345,234 |
| Né le: 04/04/2015 |
Caséine kappa: BE |
Caséine bêta: A1A2 |
|
| PRODUCTION |
| 1979 Troupeaux |
5572 Filles |
99% Fiab |
|
GEBV 25*DEC |
|
|
Lait kg
861 |
|
Gras kg
34 |
|
Gras %
0.00 |
|
Protéine kg
34 |
|
Protéine %
+0.04 |
|
Vitesse de traite
96 |
|
Tempérament
90 |
|
Efficience du méthane
95 |
|
Efficience de l'alimentation
99 |
|
BMC
98 |
|
|
|
Moyenne des filles
Lait 12,920 kg
Gras 519 kg
Protéine 420 kg
|
|
| SANTÉ ET REPRODUCTION |
|
Immunité 97 |
|
|
|
|
|
|
|
|
|
|
|
| Aptitude des filles au vêlage | 102 |
|
|
|
|
| Résistance aux maladies métaboliques | 95 |
|
|
| Résistance à la mammite | 104 |
|
|
|
|
| Persistance de lactation | 97 |
|
|
|
|
|
| CONFORMATION |
| 1865 Troupeaux |
5134 Filles |
99% Fiab |
|
GEBV 25*DEC |
|
|
|
|
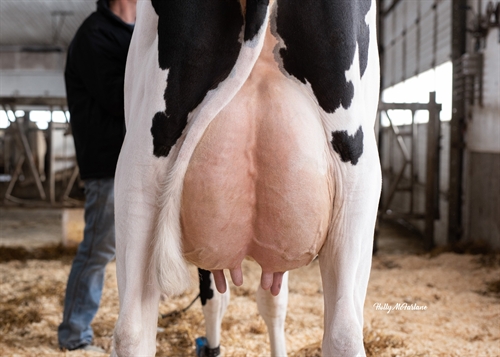
| Profondeur du pis |
|
Haut |
3H |
| Texture du pis |
|
Souple |
5 |
| Suspension médiane |
|
Forte |
5 |
| Attache avant |
|
Faible |
-1 |
| Pos. trayons avant |
|
Éloignés Ecartés |
6É |
| Haut. Arrière-pis |
|
Haute |
6 |
| Larg. Arrière-pis |
|
Large |
5 |
| Position trayons arrière |
|
Rapprochés |
2R |
| Longueur des trayons |
|
Longs |
1B |
| Angle du pied |
|
Peu profond |
0 |
| Profondeur du talon |
|
Profond |
1 |
| Qualité de l'ossature |
|
Grossière |
0 |
| Mbres arr. Vue côté |
|
Courbés |
2C |
| Mbres arr. Vue arr. |
|
Jarrets droits |
10 |
| Stature |
|
Grande |
5G |
| Largeur du poitrail |
|
Étroit |
-2 |
| Profondeur de corps |
|
Peu profond |
-8 |
| Structure des côtes |
|
Non angulaire |
0 |
| Force du rein |
|
Fort |
7 |
| Angle de la croupe |
|
Isch. bas |
2B |
| Largeur aux ischions |
|
Large |
8 |
|
|
|
| |
|
|
|
|