|
|
 |
| MAVERICK-I CRUSH |
|
| OH-RIVER-SYC MOGAL BRYLAN VG-87-4YR-USA |
|
|
|
|
| MOUNTFIELD SSI DCY MOGUL |
|
| OH-RIVER-SYC MRDN BILLIE EX-91-4YR-USA |
|
|
|
|
| SULLY HART MERIDIAN |
|
| SANDY-VALLEY ATWD BARBIE EX-91-5YR-USA |
|
|
|
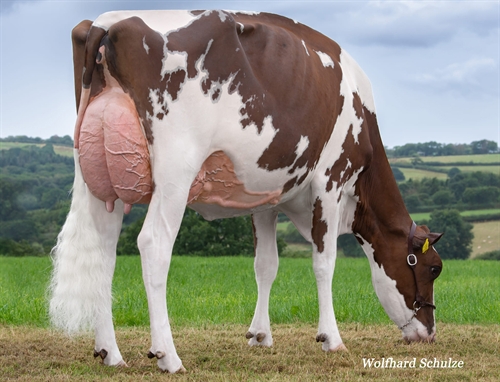
| EX-92-CAN TD TR TL TY MWT TV 99%-I |
HH1F HH2F HH3F HH4F HH5C HH6F HCDF HMWF |
| Reg. #: HO840M3135145001 |
aAa: 324165 |
DMS: 126 |
| Nacimiento: 03/10/2017 |
Caseina Kappa: BE |
Caseina Beta: A1A2 |
|
| PRODUCCIÓN |
| 3981 Hatos |
12474 Hijas |
99% Conf. |
|
MACE-G / 04-26 |
|
|
Leche lbs
-159 |
|
Grasa lbs
-61 |
|
Grasa %
-0.21 |
|
Proteína lbs
-19 |
|
Proteína %
-0.05 |
|
| NM$ -601 |
|
CM$ -621 |
|
FM$ -554 |
|
GM$ -579 |
|
DWP$ -720 |
|
Eficiencia de Conversión
-147 |
|
IR
19 |
|
Comida Ahorrada
-5 |
|
Eficiencia Metano
104 |
|
Rapidez en el Ordeñe
6.90 |
|
|
Promedio de la Hija (kg-ME)
Leche 25,570 lbs
Grasa 1,000 lbs
Proteína 824 lbs
|
|
| SALUD Y FERTILIDAD |
|
Inmunidad 92 |
|
|
|
| Inmunidad de los terneros | 106 | |
|
|
|
|
|
|
| Tasa de preñez de las hijas | -1.3 |
|
|
|
|
|
|
| Facilidad del Parto | 2.2% | 98% Conf. |
|
|
|
|
| Facilidad del Parto - Hijas | 2.2% | 96% Conf. |
|
|
|
|
|
|
|
|
| Hijas con crías muertas | 6.1% | |
|
|
|
|
| CONFORMACIÓN |
| 2807 Hatos |
8205 Hijas |
99% Conf. |
|
MACE / 04-26 |
|
|
|
|
|
|
|
|
|
| Compuesto de Patas y Pezuñas | 0.58 |
|
|
|
|
|
|
|
| Estatura |
|
|
Alta |
+2.36 |
| Fortaleza |
|
|
Fuerte |
+0.37 |
| Prof. Corporal |
|
|
Profunda |
+1.79 |
| Forma Lechera |
|
|
Angular |
+2.79 |
| A. de la Grupa |
|
|
Isquiones Bajos |
+0.39 |
| Amplitud de Grupa |
|
|
Ancha |
+1.66 |
| Vista Lat. P. Traseras |
|
|
Curvas |
+2.87 |
| Patas Traseras, Vista de Atrás |
|
|
Corvejónes Rectos |
+0.44 |
| Ang. Talón |
|
|
Alto |
+0.37 |
| Compuesto de Patas y Pezuñas |
|
|
Alto |
+1.43 |
| Inserción ubre Delantera |
|
|
Fuerte |
+2.52 |
| Ancho Ubre Trasera |
|
|
Ancha |
+2.77 |
| Altura Ubre Trasera |
|
|
Alta |
+2.07 |
| Ligamento Central |
|
|
Fuerte |
+0.98 |
| Prof. Ubre |
|
|
Poco Profundo |
+1.54 |
| Posición Pezones Delanteros |
|
|
Cerrados |
+1.53 |
| Longitud de Pezones |
|
|
Cortos |
-0.25 |
| Posición Pezones Traseros |
|
|
Cerrados |
+1.29 |
|
|
|
| |
|
|
|
|