|
|
 |
| SILVERRIDGE V EINSTEIN |
|
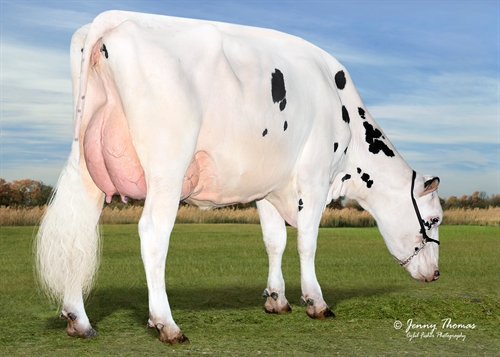
| OCD FRAZZL CAITLYN 44045 |
|
|
|
|
| MELARRY JOSUPER FRAZZLED |
|
| OCD RUBICON CAITLYN 4553 EX-90-3YR-USA DOM |
|
|
|
|
| EDG RUBICON |
|
| EDG CAITLYN DOORMAN 2274 VG-86-4YR-USA |
|
|
|
| TD TR TL TY MWT TV 99%-I |
HH1F HH2F HH3F HH4F HH5F HH6F HCDF HMWF |
| Num.Reg #: HO840M3215229448 |
aAa: 342516 |
DMS: 345,456 |
| Nacimiento: 02/27/2020 |
Kappa Caseina: AB |
Beta Caseina: A2A2 |
|
| PRODUCCION |
| 91 Rebaños |
504 Hijas |
88% Rep. |
|
MACE-G / 04-26 |
|
|
Leche lbs
239 |
|
Grasa lbs
73 |
|
Grasa %
+0.24 |
|
Proteína lbs
24 |
|
Proteína %
+0.06 |
|
| NM$ 584 |
|
CM$ 613 |
|
FM$ 518 |
|
GM$ 537 |
|
DWP$ 618 |
|
Eficiencia de Conversiòn
167 |
|
IR
-9 |
|
Comida Ahorrada
-78 |
|
Eficiencia Metano
99 |
|
Velocidad Ordeño
6.85 |
|
| SALUD Y REPRODUCCIÓN |
|
Immunity 109 |
|
|
|
|
|
|
|
|
|
| Fertilidad de las Hijas | -1.1 |
|
|
|
|
|
|
| Facilidad de Parto | 1.4% | 58% Rep. |
|
|
|
|
| Facilidad de Parto de las Hijas | 1.7% | 39% Rep. |
|
|
|
|
| Crías del Toro que Nacen Muertas | 3.6% | |
|
|
|
|
| Crías de las Hijas que Nacen Muertas | 4.5% | |
|
|
|
|
| TIPO |
| 39 Rebaños |
160 Hijas |
88% Rep. |
|
MACE / 04-26 |
|
|
|
|
| Estatura |
|
|
Baja |
-0.45 |
| Fortaleza |
|
|
Fuerte |
+0.57 |
| Profundidad Corporal |
|
|
Profunda |
+0.13 |
| Estructura Lechera |
|
|
Cost. Cerradas |
-0.12 |
| Ángulo de Grupa |
|
|
Isq. Altos |
-0.58 |
| Anchura Grupa |
|
|
Ancha |
+0.96 |
| Patas Vista Lateral |
|
|
Rectas |
-1.39 |
| Patas Vista Posterior |
|
|
Cerradas |
-0.03 |
| Ángulo Podal |
|
|
Profundo |
+0.52 |
| Colocación de las Patas |
|
|
Correcta |
+0.21 |
| Inserción Anterior |
|
|
Fuerte |
+1.26 |
| Altura Inserción Posterior |
|
|
Alta |
+1.30 |
| Anchura Inserción Posterior |
|
|
Ancha |
+1.65 |
| Ligamento Suspensor |
|
|
Fuerte |
+1.26 |
| Profundidad Ubre |
|
|
Recogida |
+1.30 |
| Colocación Pezones Anteriores |
|
|
Cerrados |
+1.29 |
| Longitud de Pezones |
|
|
Cortos |
-0.75 |
| Colocación Pezones Posteriores |
|
|
Cerrados |
+1.32 |
|
|
|
| |
|
|
|
|