|
|
 |
| CLAYNOOK BAROLO SG |
|
| DICKLANDS KING DOC 3503 VG-87-4YR-CAN 2* |
|
|
|
|
| WOODCREST KING DOC |
|
| DICKLANDS CHIEF 2630 3041 VG-85-3YR-CAN |
|
|
|
|
| STANTONS CHIEF |
|
| DICKLANDS JEDI 2630 GP-81-2YR-CAN 1* |
|
|
|
| TD TM TR TL TY MWT TV 99%-I |
HH1F HH2F HH3F HH4F HH5F HH6F HCDF HMWF |
| Num.Reg #: HOCANM14128958 |
aAa: 243516 |
DMS: 126,246 |
| Nacimiento: 08/15/2021 |
Kappa Caseina: BB |
Beta Caseina: A2A2 |
|
| PRODUCCION |
| G Rebaños |
G Hijas |
82% Rep. |
|
CDCB-G / 04-26 |
|
|
Leche lbs
262 |
|
Grasa lbs
22 |
|
Grasa %
+0.04 |
|
Proteína lbs
15 |
|
Proteína %
+0.02 |
|
| NM$ 5 |
|
CM$ 13 |
|
FM$ -16 |
|
GM$ -30 |
|
DWP$ -129 |
|
Eficiencia de Conversiòn
57 |
|
IR
-34 |
|
Comida Ahorrada
-70 |
|
Eficiencia Metano
103 |
|
Velocidad Ordeño
6.81 |
|
| SALUD Y REPRODUCCIÓN |
|
Immunity 93 |
|
|
|
|
|
|
|
|
|
| Fertilidad de las Hijas | -2.7 |
|
|
|
|
|
|
| Facilidad de Parto | 2.0% | 74% Rep. |
|
|
|
|
| Facilidad de Parto de las Hijas | 2.5% | 70% Rep. |
|
|
|
|
| Crías del Toro que Nacen Muertas | 4.7% | |
|
|
|
|
| Crías de las Hijas que Nacen Muertas | 5.7% | |
|
|
|
|
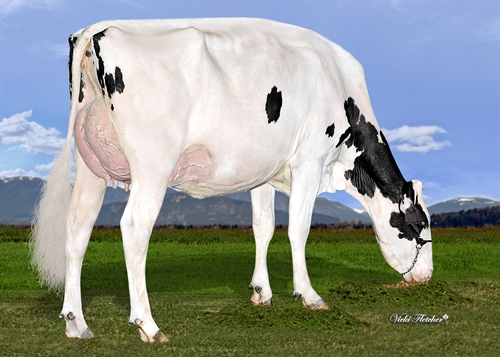
| TIPO |
| G Rebaños |
G Hijas |
81% Rep. |
|
HAUSA-G / 04-26 |
|
|
|
|
| Estatura |
|
|
Alta |
+2.23 |
| Fortaleza |
|
|
Fuerte |
+1.12 |
| Profundidad Corporal |
|
|
Profunda |
+1.74 |
| Estructura Lechera |
|
|
Cost. Abierta |
+2.45 |
| Ángulo de Grupa |
|
|
Isq. Altos |
-0.47 |
| Anchura Grupa |
|
|
Ancha |
+1.96 |
| Patas Vista Lateral |
|
|
Curvas |
+1.58 |
| Patas Vista Posterior |
|
|
Aplomadas |
+1.18 |
| Ángulo Podal |
|
|
Profundo |
+1.13 |
| Colocación de las Patas |
|
|
Correcta |
+1.31 |
| Inserción Anterior |
|
|
Fuerte |
+1.67 |
| Altura Inserción Posterior |
|
|
Alta |
+1.49 |
| Anchura Inserción Posterior |
|
|
Ancha |
+1.69 |
| Ligamento Suspensor |
|
|
Fuerte |
+1.79 |
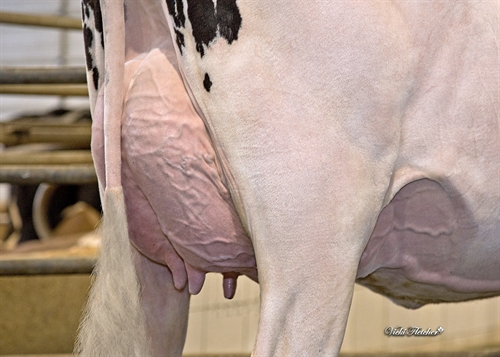
| Profundidad Ubre |
|
|
Recogida |
+1.21 |
| Colocación Pezones Anteriores |
|
|
Cerrados |
+1.45 |
| Longitud de Pezones |
|
|
Cortos |
-0.67 |
| Colocación Pezones Posteriores |
|
|
Cerrados |
+1.28 |
|
|
|
| |
|
|
|
|