|
|
 |
| PEAK ZAZZLE |
|
| PROGENESIS HOT VACATION VG-86-4YR-CAN 1* |
|
|
|
|
| PEAK HOTJOB |
|
| FB LULU ACHV VEGAS EX-90-2E-CAN 2* |
|
|
|
|
| ABS ACHIEVER |
|
| STUNNING-M MONTROSS LULU VG-87-2YR-USA DOM |
|
|
|
| GLPI +3587 PRO$ 2476 |
| DPF RDF BLF CNF BYF MWF CVF |
HH1F HH2F HH3F HH4F HH5F HH6C HCDF HMWF |
| Reg. #: HOCANM13807684 |
aAa: 243615 |
DMS: 126,561 |
| Data nascita: 07/06/2020 |
Kappa Caseina: AB |
Beta Caseina: A1A2 |
|
| PRODUZIONE |
| 76 Allev. |
171 Figlie |
95% Att. |
|
GEBV 26*APR |
|
|
Latte kg
-514 |
|
Grasso kg
104 |
|
Grasso %
+1.06 |
|
Proteina kg
34 |
|
Proteina %
+0.41 |
|
Velocità di mungitura
103 |
|
Temperamento
101 |
|
Efficienza Metano
106 |
|
Efficienza Alimentare
98 |
|
Condizione Corporea
103 |
|
|
|
EVM Figlie in Canada
Latte 12,488 kg
Grasso 578 kg
Proteina 433 kg
|
|
| TRATTI GESTIONALI |
|
Immunity 101 |
|
|
|
|
|
|
|
|
|
| Fertilità delle figlie | 108 |
|
|
| Facilità al parto delle figlie | 106 |
|
|
| Punteggio Condizione Corporea | 102 |
|
|
| Resistenza Malattie Metaboliche | 103 |
|
|
| Resistenza alle Mastiti | 104 |
|
|
|
|
| Persistenza Lattazione | 105 |
|
|
|
|
|
| MORFOLOGIA |
| 68 Allev. |
154 Figlie |
92% Att. |
|
GEBV 26*APR |
|
|
|
|
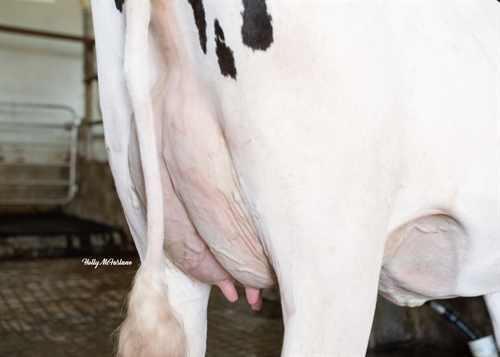
| Profondità Mammella |
|
Alta |
2A |
| Qualità del Tessuto |
|
Carnoso |
-1 |
| Legamento |
|
Debole |
-6 |
| Mammella Anteriore |
|
Forte |
6 |
| Posiz. Capezzoli ant. |
|
Convergenti |
1C |
| Mamm. Post. Altezza |
|
Alta |
1 |
| Mamm. Post. Larghezza |
|
Stretta |
-3 |
| Posiz. Capezzoli post. |
|
Convergenti |
1C |
| Lunghezza Capezzoli |
|
Lunghi |
1L |
| Angolo del Piede |
|
Alto |
4 |
| Altezza Tallone |
|
Alto |
6 |
| Qualitá dell' Ossatura |
|
Fine |
6 |
| Arti post. Vista lat. |
|
Stangati |
3S |
| Arti post. Vista post. |
|
In appiombo |
2 |
| Statura |
|
Alta |
1A |
| Larghezza torace |
|
Stretta |
-4 |
| Profondità |
|
Poco profondo |
-8 |
| Angolosità |
|
Rotonda |
-5 |
| Forza dei Lombi |
|
Forte |
3 |
| Angolo Groppa |
|
Spiovente |
4S |
| Larghezza agli Ischi |
|
Larga |
2 |
|
|
|
| |
|
|
|
|