|
|
 |
| DYMENTHOLM S SYMPATICO *RC |
|
| VAL-BISSON SHOTTLE IMELDA EX-91-6YR-CAN 11* |
|
|
|
|
| PICSTON SHOTTLE |
|
| VAL-BISSON GOLDWYN MAYA VG-88-6YR-CAN 42* |
|
|
|
|
| BRAEDALE GOLDWYN |
|
| VAL-BISSON FINLEY DREAM VG-87-6YR-CAN 14* |
|
|
|
| GP-CAN TD RC TL TY TV |
HH1F HH2F HH3C HH4F HH5F HH6F HCDF HMWF |
| Rek #: HOCANM108109390 |
aAa: 312546 |
DMS: 135 |
| Syntynyt: 05/18/2013 |
Kappakaseiini: AB |
Beettakaseiini: A1A2 |
|
| TUOTOS |
| 1603 Karjoja |
3493 Tytt |
99% arv.varm |
|
MACE-G / 12-25 |
|
|
Maito paunaa
-1590 |
|
Rasva paunaa
-13 |
|
Rasva %
+0.21 |
|
Valkuainen paunaa
-46 |
|
Valkuainen %
+0.02 |
|
| NM$ -593 |
|
CM$ -586 |
|
FM$ -607 |
|
GM$ -599 |
|
DWP$ -750 |
|
Rehuhyötysuhde
-168 |
|
RFI
295 |
|
Rehunsäästö
-519 |
|
Metaanitehokkuus
96 |
|
Lypsynopeus
6.77 |
|
|
Tytärten aikuistuotos (ME)
Maito 23,377 paunaa
Rasva 974 paunaa
Valkuainen 752 paunaa
|
|
| TERVEYS JA HEDELMÄLLISYYS |
|
Immuunivaste 90 |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
| Poikimavaikeus isänä | 1.7% | 96% arv.varm |
|
|
|
|
| Poikimavaikeus emänisänä | 3.4% | 93% arv.varm |
|
|
| hedelmällisyysindeksi | -0.4 |
|
|
| Vasikkakuolleisuus isänä | 3.6% | |
|
|
|
|
| Vasikkakuolleisuus emänisänä | 5.1% | |
|
|
|
|
| RAKENNE |
| 1097 Karjoja |
2107 Tytt |
98% arv.varm |
|
MACE / 12-25 |
|
|
|
|
| Takakorkeus |
|
|
korkea |
+0.88 |
| Vahvuus |
|
|
vahva |
+1.00 |
| Rungon syvyys |
|
|
syvä |
+0.73 |
| Lypsytyyppisyys |
|
|
huono |
-0.55 |
| Lantiokulma |
|
|
nouseva |
-1.35 |
| Lantion leveys |
|
|
leveä |
+0.82 |
| Kinnerkulma |
|
|
suora |
-1.48 |
| Takajalat takaa |
|
|
suorat |
+1.83 |
| Sorkkakulma |
|
|
hyvä |
+0.98 |
| Jalkarakenne |
|
|
hyvä |
+0.88 |
| Etukiinnitys |
|
|
hyvä |
+1.59 |
| Takakiinnityksen korkeus |
|
|
hyvä |
+1.91 |
| Takakiinnityksen leveys |
|
|
leveä |
+0.67 |
| Keskiside |
|
|
vahva |
+0.84 |
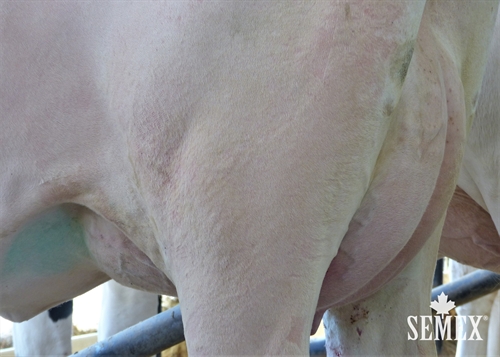
| Utaremuoto |
|
|
laaka |
+1.77 |
| Etuvedinten sijainti |
|
|
lähellä |
+0.84 |
| Etuvedinten pituus |
|
|
lyhyet |
-1.07 |
| Takavedinten sijainti |
|
|
lähellä |
+1.21 |
|
|
|
| |
|
|
|
|