|
|
 |
| SILVERRIDGE V TIMBERLAKE |
|
| SIEMERS TRJ BROOKE 28030 GP-84-2YR-USA |
|
|
|
|
| SIEMERS TRAJECTORY |
|
| SIEMERS DELTA BROOKE-D7 GP-81-3YR-USA |
|
|
|
|
| MR MOGUL DELTA 1427 |
|
| MAPEL WOOD MAN O MAN BROOKE VG-86-2YR-CAN 4* |
|
|
|
| GLPI +3075 PRO$ 1675 |
| DPF RDF BLF CNF BYF MWF CVF |
HH1F HH2F HH3F HH4F HH5F HH6F HCDC HMWF |
| Reg. #: HO840M3143986627 |
aAa: 423651 |
DMS: 345,135 |
| Data nascita: 06/14/2019 |
Kappa Caseina: AB |
Beta Caseina: A2A2 |
|
| PRODUZIONE |
| 39 Allev. |
195 Figlie |
96% Att. |
|
GEBV 26*APR |
|
|
Latte kg
803 |
|
Grasso kg
32 |
|
Grasso %
0.00 |
|
Proteina kg
63 |
|
Proteina %
+0.27 |
|
Velocità di mungitura
101 |
|
Temperamento
106 |
|
Efficienza Metano
87 |
|
Efficienza Alimentare
102 |
|
Condizione Corporea
106 |
|
|
|
EVM Figlie in Canada
Latte 13,601 kg
Grasso 553 kg
Proteina 464 kg
|
|
| TRATTI GESTIONALI |
|
Immunity 99 |
|
|
|
|
|
|
|
|
|
|
|
| Facilità al parto delle figlie | 103 |
|
|
| Punteggio Condizione Corporea | 92 |
|
|
| Resistenza Malattie Metaboliche | 101 |
|
|
| Resistenza alle Mastiti | 102 |
|
|
|
|
| Persistenza Lattazione | 102 |
|
|
|
|
|
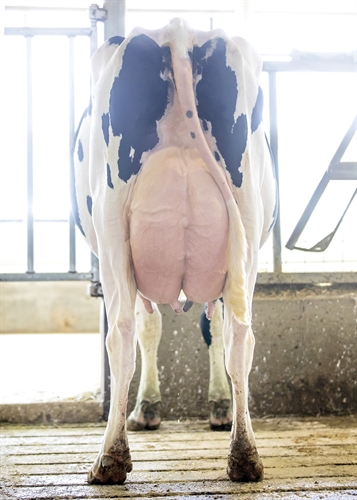
| MORFOLOGIA |
| 29 Allev. |
84 Figlie |
91% Att. |
|
GEBV 26*APR |
|
|
|
|
| Profondità Mammella |
|
Alta |
5A |
| Qualità del Tessuto |
|
Carnoso |
-1 |
| Legamento |
|
Debole |
-5 |
| Mammella Anteriore |
|
Forte |
4 |
| Posiz. Capezzoli ant. |
|
Divergenti |
6D |
| Mamm. Post. Altezza |
|
Alta |
7 |
| Mamm. Post. Larghezza |
|
Larga |
2 |
| Posiz. Capezzoli post. |
|
Divergenti |
5D |
| Lunghezza Capezzoli |
|
Lunghi |
4L |
| Angolo del Piede |
|
Basso |
-7 |
| Altezza Tallone |
|
Alto |
1 |
| Qualitá dell' Ossatura |
|
Grosso |
-1 |
| Arti post. Vista lat. |
|
Falciati |
9F |
| Arti post. Vista post. |
|
Vaccini |
-11 |
| Statura |
|
Bassa |
7B |
| Larghezza torace |
|
Stretta |
-9 |
| Profondità |
|
Poco profondo |
-10 |
| Angolosità |
|
Rotonda |
-1 |
| Forza dei Lombi |
|
Debole |
-5 |
| Angolo Groppa |
|
Alto |
0 |
| Larghezza agli Ischi |
|
Stretta |
-7 |
|
|
|
| |
|
|
|
|