|
|
 |
| CLAYNOOK ZASBERILLA |
|
| PROGENESIS HOT VACATION VG-86-4YR-CAN 1* |
|
|
|
|
| PEAK HOTJOB |
|
| FB LULU ACHV VEGAS EX-90-2E-CAN 2* |
|
|
|
|
| ABS ACHIEVER |
|
| STUNNING-M MONTROSS LULU VG-87-2YR-USA DOM |
|
|
|
| TD TR TL TY MWT TV 99%-I |
HH1F HH2F HH3F HH4F HH5F HH6F HCDF HMWF |
| Rek #: HOCANM13807972 |
aAa: 243651 |
DMS: 561,456 |
| Syntynyt: 10/15/2020 |
Kappakaseiini: AB |
Beettakaseiini: A2A2 |
|
| TUOTOS |
| 13 Karjoja |
273 Tytt |
96% arv.varm |
|
CDCB-G / 12-25 |
|
|
Maito paunaa
75 |
|
Rasva paunaa
67 |
|
Rasva %
+0.24 |
|
Valkuainen paunaa
26 |
|
Valkuainen %
+0.09 |
|
| NM$ 450 |
|
CM$ 489 |
|
FM$ 363 |
|
GM$ 407 |
|
DWP$ 591 |
|
Rehuhyötysuhde
149 |
|
RFI
33 |
|
Rehunsäästö
-159 |
|
Metaanitehokkuus
101 |
|
Lypsynopeus
7.36 |
|
|
Tytärten aikuistuotos (ME)
Maito 27,682 paunaa
Rasva 1,240 paunaa
Valkuainen 952 paunaa
|
|
| TERVEYS JA HEDELMÄLLISYYS |
|
Immuunivaste 107 |
|
|
|
|
|
|
|
|
|
|
|
| Hiehojen tiinehtyvyys | -0.2 | |
|
|
|
|
| Poikimavaikeus isänä | 1.0% | 74% arv.varm |
|
|
|
|
| Poikimavaikeus emänisänä | 1.3% | 72% arv.varm |
|
|
| hedelmällisyysindeksi | -1.0 |
|
|
| Vasikkakuolleisuus isänä | 3.5% | |
|
|
|
|
| Vasikkakuolleisuus emänisänä | 3.5% | |
|
|
|
|
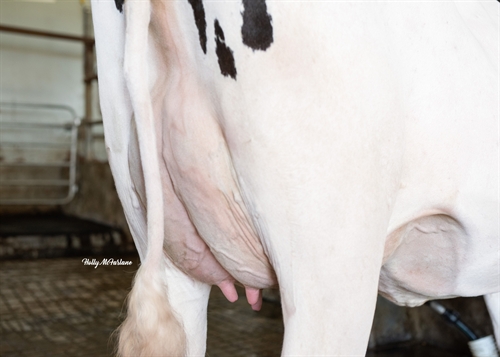
| RAKENNE |
| 4 Karjoja |
96 Tytt |
84% arv.varm |
|
HAUSA-G / 12-25 |
|
|
|
|
| Takakorkeus |
|
|
korkea |
+0.12 |
| Vahvuus |
|
|
vahva |
+0.46 |
| Rungon syvyys |
|
|
huono |
-0.16 |
| Lypsytyyppisyys |
|
|
huono |
-0.60 |
| Lantiokulma |
|
|
nouseva |
-1.19 |
| Lantion leveys |
|
|
leveä |
+0.88 |
| Kinnerkulma |
|
|
suora |
-0.62 |
| Takajalat takaa |
|
|
pihtiset |
-0.86 |
| Sorkkakulma |
|
|
hyvä |
+0.27 |
| Jalkarakenne |
|
|
hyvä |
+0.06 |
| Etukiinnitys |
|
|
hyvä |
+1.70 |
| Takakiinnityksen korkeus |
|
|
hyvä |
+1.28 |
| Takakiinnityksen leveys |
|
|
leveä |
+1.17 |
| Keskiside |
|
|
vahva |
+0.69 |
| Utaremuoto |
|
|
laaka |
+1.35 |
| Etuvedinten sijainti |
|
|
lähellä |
+0.24 |
| Etuvedinten pituus |
|
|
lyhyet |
-0.12 |
| Takavedinten sijainti |
|
|
lähellä |
+0.53 |
|
|
|
| |
|
|
|
|