|
|
 |
| PROGENESIS MONTEVERDI |
|
| WESTCOAST FSTBL FAR8167 10707 VG-85-3YR-CAN |
|
|
|
|
| BOMAZ FASTBALL |
|
| WESTCOAST CHLNGR FAR6156 8167 EX-90-2E-CAN 3* |
|
|
|
|
| SANDY-VALLEY CHALLENGER |
|
| WESTCOAST IMAX FARIA 6156 2* |
|
|
|
| TD TM TR TL TY MWT TV 99%-I |
HH1F HH2F HH3F HH4F HH5F HH6C HCDF HMWF |
| Reg. #: HOCANM14219829 |
aAa: 423651 |
DMS: 135,561 |
| Data nascita: 01/21/2022 |
Kappa Caseina: AB |
Beta Caseina: A1A2 |
|
| PRODUZIONE |
| G Allev. |
G Figlie |
82% Att. |
|
CDCB-G / 04-26 |
|
|
Latte lbs
1984 |
|
Grasso lbs
72 |
|
Grasso %
-0.04 |
|
Proteina lbs
64 |
|
Proteina %
0.00 |
|
| NM$ 842 |
|
CM$ 849 |
|
FM$ 826 |
|
GM$ 809 |
|
DWP$ 722 |
|
Efficienza Alimentare
277 |
|
RFI
-139 |
|
Risparmio Sostanza Secca
274 |
|
Efficienza Metano
97 |
|
Velocità di mungitura
7.35 |
|
| TRATTI GESTIONALI |
|
Immunity 97 |
|
|
|
|
|
|
|
| Tasso Concepimento Vacche | -0.8 | |
|
|
|
|
| Tasso Concepimento Manze | 0.5 | |
|
|
|
|
| Facilità al parto | 1.1% | 83% Att. |
|
|
| Ind. Sopravvivenza Manze | 0.6 |
|
|
| Facilità al parto delle figlie | 1.6% | 71% Att. |
|
|
|
|
|
|
|
|
| Vitalità figli delle figlie | 4.0% | |
|
|
|
|
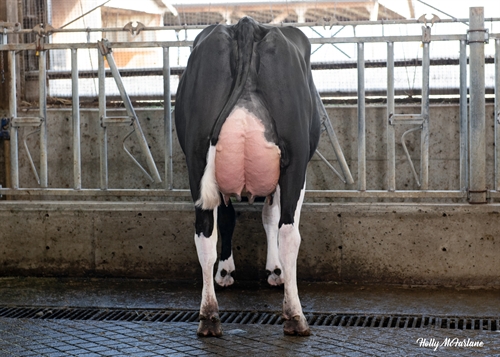
| MORFOLOGIA |
| G Allev. |
G Figlie |
81% Att. |
|
HAUSA-G / 04-26 |
|
|
|
|
| Statura |
|
|
Alta |
+0.75 |
| Forza-Vigore |
|
|
Debole |
-0.50 |
| Profondità |
|
|
Poco profondo |
-0.13 |
| Caratteri da Latte |
|
|
Cost. Aperto |
+1.49 |
| Angolo Groppa |
|
|
Spiovente |
+1.99 |
| Groppa larghezza |
|
|
Larga |
+0.40 |
| Arti post. Vista lat. |
|
|
Stangati |
-1.15 |
| Arti post. Vista post. |
|
|
Vaccini |
-0.74 |
| Angolo del Piede |
|
|
Basso |
-0.11 |
| Mobilità |
|
|
Non desiderabile |
-0.56 |
| Mammella Anteriore |
|
|
Forte |
+0.94 |
| Mamm. Post. Altezza |
|
|
Alta |
+1.67 |
| Mamm. Post. Larghezza |
|
|
Larga |
+1.49 |
| Legamento |
|
|
Forte |
+0.14 |
| Profondità Mammella |
|
|
Alta |
+0.44 |
| Posiz. Capezzoli ant. |
|
|
Divergenti |
-0.10 |
| Capezzoli lunghezza |
|
|
Corti |
-0.37 |
| Posiz. Capezzoli post. |
|
|
Divergenti |
-0.01 |
|
|
|
| |
|
|
|
|