|
|
 |
| STANTONS COCKPIT |
|
| SILVERRIDGE V MARIUS EVICTION EX-91-5YR-CAN 4* |
|
|
|
|
| PROGENESIS MARIUS |
|
| SILVERRIDGE V DUKE EMAIL EX-90-3E-CAN 11* |
|
|
|
|
| S-S-I MONTROSS DUKE |
|
| SILVERRIDGE V SSHOT EARRING EX-90-6YR-CAN 11* |
|
|
|
| TD TR TL TY MWT TV 99%-I |
HH1F HH2F HH3F HH4F HH5F HH6F HCDF HMWF |
| Nom #: HOCANM13848151 |
aAa: 243651 |
DMS: 126,246 |
| Né le: 11/24/2020 |
Caséine kappa: AB |
Caséine bêta: A1A2 |
|
| PRODUCTION |
| 50 Troup |
133 Filles |
92% Fiab |
|
MACE-G / 04-26 |
|
|
Lait lbs
1110 |
|
Gras lbs
69 |
|
Gras %
+0.08 |
|
Protéine lbs
38 |
|
Protéine %
+0.01 |
|
| NM$ 530 |
|
CM$ 541 |
|
FM$ 508 |
|
GM$ 475 |
|
DWP$ 568 |
|
Efficience de l'alimentation
188 |
|
IAR
42 |
|
Alimentation économisée
-28 |
|
Efficience du méthane
101 |
|
Vitesse de traite
6.81 |
|
|
Moyenne des filles
Lait 26,951 lbs
Gras 1,127 lbs
Protéine 895 lbs
|
|
| SANTÉ ET REPRODUCTION |
|
Immunité 106 |
|
|
|
|
|
|
|
| Taux de conception des vaches | -1.6 | |
|
|
|
|
| Taux de conception des génisses | -2.5 | |
|
|
|
|
| Facilité de vêlage | 1.6% | 92% Fiab |
|
|
| Livabilité des génisses | -0.8 |
|
|
| Facilité vêl. Filles | 1.7% | 82% Fiab |
|
|
|
|
|
|
|
|
| Mortinatalité des filles | 4.6% | |
|
|
|
|
| CONFORMATION |
| 21 Troup |
35 Filles |
87% Fiab |
|
MACE / 04-26 |
|
|
|
|
| Stature |
|
|
Grande |
+0.76 |
| Force |
|
|
Forte |
+0.42 |
| Profondeur de corps |
|
|
Profond |
+0.76 |
| Car. Laitier |
|
|
Côtes ouvertes |
+1.70 |
| Angle de la croupe |
|
|
Isch. bas |
+1.19 |
| Largeur de la croupe |
|
|
Large |
+1.06 |
| Mbres arr. Vue côté |
|
|
Courbés |
+0.32 |
| Mbres arr. Vue arr. |
|
|
Jarrets serrés |
-0.92 |
| Angle du pied |
|
|
Profond |
+0.32 |
| Indice pieds & mbres |
|
|
Indésirable |
-0.21 |
| Attache avant |
|
|
Forte |
+1.50 |
| Haut. Arrière-pis |
|
|
Haute |
+1.46 |
| Larg. Arrière-pis |
|
|
Large |
+2.37 |
| Suspension médiane |
|
|
Faible |
0.00 |
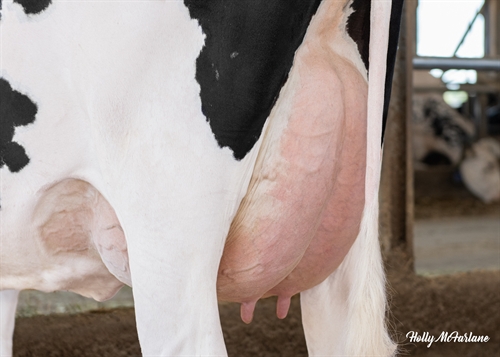
| Profondeur du pis |
|
|
Haut |
+0.57 |
| Pos. trayons avant |
|
|
Rapprochés |
+0.93 |
| Longueur des trayons |
|
|
Courts |
-1.03 |
| Position trayons arrière |
|
|
Rapprochés |
+0.34 |
|
|
|
| |
|
|
|
|